孟德尔两大定律的机制
随着生殖细胞成熟过程接近尾声,同等大小的染色体接合成对。接着,细胞分裂,每对染色体中的两条各自进入一个细胞。每个成熟的生殖细胞将只包含一组染色体。(图3-1和图3-2)
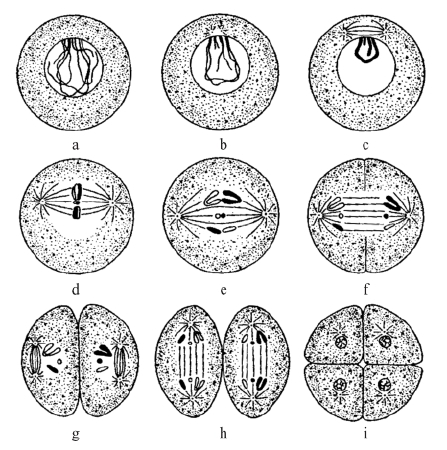
图 3-1 精细胞的成熟
注:图示精细胞的两次成熟分裂。每个细胞表现三对染色体;黑色代表来自父方的染色体,白色代表来自母方的染色体(a、b、c除外)。图d、e、f所示的第一次成熟分裂为减数分裂。图g、h所示的第二次成熟分裂为均等分裂,每条染色体纵裂为两条新染色体。

图 3-2 卵细胞的成熟
注:图示卵细胞的两次成熟分裂。第一次成熟分裂的纺锤体如图a所示;父方染色体与母方染色体的分离(减数分裂)如图b所示;第一极体分出如图c所示;第二次成熟分裂的纺锤体如图d所示,每条染色体纵裂为两条新染色体(均等分裂);第二极体形成如图e所示;卵细胞核只有一半子染色体(单倍体)如图f所示。
在成熟阶段,染色体的这种行为与孟德尔第一定律相似,每对染色体中,来自父方的染色体与来自母方的染色体分离。结果生殖细胞包含每对染色体中的一条。关于每一对染色体,我们可以指出,在它成熟后,一半生殖细胞包含每对染色体中的一条,另一半则含有这对染色体中的另一条。如果把染色体换成孟德尔单元,其表述是相同的。
每对染色体中有一条来自父方,另一条来自母方。如果成对接合的染色体在纺锤体上排列时,所有来自父方的染色体都聚集在一端,所有来自母方的染色体都聚集在另一端,这样产生的两个生殖细胞将分别与父亲或母亲的生殖细胞相同。我们没有一个先验的理由来假定接合的染色体将按照这一方式运作,但想要证明它们没有这样做却是极端困难的,因为就这一事例的本质而言,正在接合中的两条染色体在形状与尺寸上都类似,没有哪一规则可以通过观察告诉我们哪一条染色体是父方的,哪一条又是母方的。
不过,近年来,在一些对蚱蜢进行研究的例子中,发现某些成对的染色体在形状或纺锤体纤维的联系方式上有时存在微弱差别。(图3-3)当生殖细胞成熟时,这些染色体先两两接合,然后分离。由于它们保留着个体差异,所以能够追踪到它们进入两极的踪迹。
现在,在这些蚱蜢里,雄性有一条不成对的染色体,与雄性性别的决定机制有关。(图3-3)在成熟分裂时,这条染色体要么进入纺锤体的这一极,要么进入纺锤体的另一极。它可以作为成对染色体行动方向的标记。卡罗瑟斯(Carothers)女士首先完成了这些观察。她看到一对具有标记性(一条弯的,一条直的)的染色体,其根据与性染色体的关系,可以向任意一极分离。
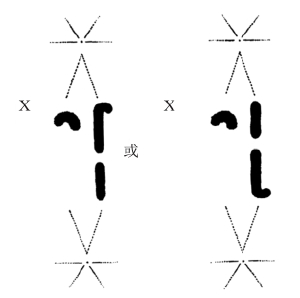
图 3-3 X染色体与一对常染色体的自由组合
就这个例子进一步观察,我们发现一些个体中的其他几对染色体有时也表现出一定的差异。一项关于这些染色体在减数分裂中的情形研究,再一次表明一对染色体向两极分布的方向与其他对染色体的分布方向互不相关。从这里我们得到了不同对染色体之间自由组合的客观证据。这一证据与孟德尔关于不同连锁群中的基因自由分配的第二定律相似。

